

Revista Iberoamericana de Neuropsicología
Vol. 1, No. 2: 180-191, julio-diciembre 2018.
Un reloj interno: modelos, neurociencias y Trastorno por Déficit de Atención e Hiperactividad
Suárez Isabel1, Moreno Torres Mayilín1, Casini Laurence2
1 Grupo de Psicología de la Universidad del Norte, Barranquilla, Colombia
2 CNRS, Laboratoire de Neurosciences Cognitives, Aix-Marseille Université, Marseille, France
Correspondence should be directed to
Dr. Isabel C. Suárez Del Chiaro.
Departamento de Psicología,
Universidad del Norte,
Km 5 vía Puerto Colombia,
Barranquilla, Colombia.
E-mail: delchiaroi@uninorte.edu.co
Phone +57 (5) 3509852
The authors have declared they have no financial interests to disclose.
Los autores han declarado que no tienen conflicto de intereses.
Este trabajo ha sido financiado por el Departamento de Investigación Desarrollo e Innovación (DIDI) de la Universidad del Norte, Barranquilla.
[su_row][su_column size=”1/2″][su_button url=”https://neuropsychologylearning.com/wp-content/uploads/pdf/pdf-revista-vol1/RevNeuro2-4-Reloj.pdf” target=”blank” style=”flat” background=”#F38546″ color=”#ffffff” size=”6″ radius=”0″ icon=”icon: arrow-down” icon_color=”#ffffff” text_shadow=”0px 0px 0px #ffffff”]ARTÍCULO[/su_button][/su_column] [su_column size=”1/2″][su_button url=”https://neuropsychologylearning.com/wp-content/uploads/pdf/pdf-revista-vol1/revis-neuro-2-160718.pdf” target=”blank” style=”flat” background=”#F38546″ color=”#ffffff” size=”6″ radius=”0″ icon=”icon: arrow-down” icon_color=”#ffffff” text_shadow=”0px 0px 0px #ffffff”]REVISTA VOL1-2[/su_button][/su_column] [/su_row]
Un reloj interno: modelos, neurociencias y Trastorno por Déficit de Atención e Hiperactividad
There are three purposes to this theoretical review. First, to present the main cognitive models of time perception based on the idea of an internal clock, as a core mechanism for making temporal judgments. Secondly, presenting the studies of cognitive neuroscience that provide evidence for cortical and subcortical structures involved this models. Specifically, we will focus on the role of the supplementary motor area (SMA) as an accumulator. Finally, we propose this model as a new framework to better understand the deficit in temporal processing in patients diagnosed with attention deficit and hyperactivity disorder.
Este artículo de revisión teórica tiene tres objetivos. Primero, presentar una revisión de los principales modelos cognitivos de la percepción del tiempo basados en la idea de la existencia de un reloj interno como mecanismo necesario para la estimación de juicios temporales. Segundo, presentar los estudios de neurociencias cognitivas que aportan evidencia de las estructuras corticales y subcorticales que participan en el funcionamiento de este reloj. En particular, el rol del área motora suplementaria (AMS) en la acumulación del tiempo. Finalmente, proponer el modelo del reloj interno, como una alternativa para una mejor comprensión del déficit en el procesamiento temporal en pacientes diagnosticados con déficit por trastorno de atención con hiperactividad (TDAH).
La estimación correcta del tiempo, es una capacidad esencial para la interacción con el entorno. Esta capacidad sirve de base para la adaptación de todas las actividades cotidianas y ayuda a anticipar los comportamientos adecuados para la ejecución de una acción y evitar sufrir accidentes. Por ejemplo, cuando un conductor se desplaza en su carro a través de una avenida congestionada, debe estimar la duración de los eventos que suceden en el camino (ej. cambio de luces de un semáforo, momento oportuno para ingresar a la autopista, entre otros). Sin embargo ¿cómo es posible que construyamos esta representación subjetiva del tiempo?
En las últimas décadas, la idea del tiempo subjetivo ha dejado de ser una noción filosófica, para constituirse en un constructo cognitivo y uno de los principales temas de investigación en psicología. El presente artículo, pretende hacer una revisión de los modelos actuales de la psicología de la percepción del tiempo, que se fundamentan en la idea de un reloj o cronómetro interno, como base de los cálculos mentales necesarios a la estimación correcta del tiempo. Además, realiza un recorrido por los principales estudios de neurociencias cognitivas en los que se evidencia el rol de una red de estructuras corticales y subcorticales implicadas en el funcionamiento de este reloj. Finalmente, proponemos este modelo, como una aproximación teórica que puede ayudar a tener una mejor comprensión del déficit en el procesamiento temporal de las personas diagnosticadas con déficit por trastorno de atención con hiperactividad (TDAH).
Históricamente, los estudios sobre la estimación del tiempo y sus bases fisiológicas comenzaron en Francia alrededor de 1927 con el investigador François Marcel (1). Sus estudios, se basaron en la idea según la cual el tiempo subjetivo dependería de un mecanismo psicofisiológico. Clásicamente, era conocido que cuando los fenómenos fisicoquímicos son sometidos a altas temperaturas, sufrían una aceleración. Según esta premisa, si el tiempo subjetivo respondía a propiedades fisicoquímicas, entonces tendría que someterse a reglas similares que rigen estos fenómenos. Para comprobar esta hipótesis, François, realizó una serie de experimentos en los que manipulaba la temperatura corporal y observaba el efecto que se producía en la estimación del tiempo de las personas. A través de la técnica diatérmica (corriente eléctrica sin peligro), midió la estimación subjetiva del tiempo en tareas de producción y de discriminación temporal.
En la prueba de producción temporal, los participantes debían producir un ritmo de 3 golpes por segundo. En cambio, en la segunda prueba, de discriminación, los participantes debían estimar la duración de los latidos presentados por un metrónomo y detenerlo después de estimar una duración de 4 latidos. Los resultados de estas experiencias mostraron una clara relación entre la temperatura corporal y la aceleración del tiempo de producción de los golpes rítmicos.
Posteriormente, el investigador inglés Hoagland se interesó en 1933 en el efecto de la temperatura sobre la estimación del tiempo, a partir de una anecdótica historia. Durante un periodo, su esposa, que sufría de fuertes fiebres, le expresó tener el sentimiento de que el tiempo transcurría más lento. Hoagland, decidió examinar si el tiempo subjetivo de su esposa había sido alterado por la enfermedad. Así pues, la sometió a una serie de pruebas de producción de tiempo. En estas pruebas, su esposa debía contar hasta 60, a un ritmo estimado de 1 golpe por segundo. Los resultados mostraron que la señora de Hoagland, aceleraba el cálculo por segundo cuando la fiebre aumentaba su temperatura corporal, es decir, se aceleraba (2) .
Años más tarde, Hoagland, descubrió los estudios llevados a cabo por François, y los integró con los que había realizado, para calcular ‘la energía de activación del reloj químico’. Los trabajos de ambos investigadores, parecían responder bien a la misma teoría de energía de activación, lo que le permitió concluir que la existencia de un proceso químico relacionado al del tiempo subjetivo.
Posteriormente, Wearden & Penton-Voak (3) realizaron una revisión de todos los trabajos que, desde 1927 hasta 1933, manipularon la temperatura corporal para modular la percepción del tiempo subjetivo de las personas. Su revisión reveló que la mayoría de los estudios utilizaron diferentes y creativos métodos, como fue el caso del estudio de la percepción subjetiva del tiempo en personas enfermas de fiebre.
En casi todos los estudios, de forma sistemática, se observaba que cuando se aumentaba la temperatura corporal se aceleraba la percepción del paso del tiempo. Sin embargo, unos pocos estudios encontraron que cuando se disminuía la temperatura corporal, se reducía esta percepción.
Psicofísica de la percepción subjetiva del tiempo.
El hecho de que una manipulación física pudiera alterar la percepción del tiempo subjetivo fue, sin lugar a duda, una prueba experimental fundamental para concebir la idea de la existencia de un reloj o cronómetro interno. Sin embargo, en aquella época, los investigadores, no tenían suficientes elementos para comprender los mecanismos de ese reloj.
En los años sesenta, gracias al auge de la psicofísica, aumentó el interés por comprender la psicología del tiempo y los mecanismos a la base del funcionamiento del reloj interno. Desde ese momento los paradigmas los prospectivos y retrospectivos han sido los más utilizados para estudiar la percepción del tiempo. En el juicio temporal prospectivo, el participante sabe con anterioridad que deberá evaluar la duración de la tarea a la que será sometido (ej. “presionar un botón durante un segundo”, “escuchar dos tonos, y decir cuál es más largo”). En contraste, en los paradigmas donde el juicio temporal es “retrospectivo”, no se previene al participante sobre la dimensión temporal de la tarea (ej. preguntas inesperadas sobre el tiempo: “¿cuánto tiempo cree usted que lleva escuchando el mensaje?”). Sin embargo, los paradigmas retrospectivos, no registran la duración del tiempo, por lo que el juicio emitido parece estar basado en informaciones que no son de naturaleza temporal, por ende, menos precisos, por lo que los paradigmas prospectivos son los utilizados para estudiar la percepción subjetiva del tiempo(4).
Clásicamente, las tareas prospectivas proporcionan un excelente contexto para la aplicación de análisis psicofísicos. Por ejemplo, en una tarea de bisección temporal, que permite medir la percepción del tiempo, el participante debe aprender a diferenciar dos duraciones de tiempo extremas (ej. dos tonos auditivos, uno corto de 150 ms y uno largo de 430 ms). Después, durante la fase de prueba, las duraciones aprendidas, se presentan con algunas duraciones intermedias (ej. 150 ms, 220 ms, 290 ms, 360 ms, 430 ms). Los participantes deben categorizar todas las duraciones, bien sea como cortas o largas.
Las respuestas de los participantes son cuantificadas en función de las duraciones largas reportadas y descritas en una función sigmoidea. Este procedimiento permite calcular dos medidas psicofísicas: el umbral diferencial (Difference Limen, DL) y el punto de igualdad subjetiva (PES). El DL proporciona una medida de la precisión perceptual. Un DL largo indica una baja resolución perceptual, mientras que un DL corto sugiere una mejor precisión perceptual. Por su parte el PSE ofrece una estimación del valor que los sujetos consideran que coincide con el estímulo estándar, ya que este valor tiene la misma probabilidad de ser juzgado como mayor o menor que el de referencia.
Por otro lado, una de las leyes psicofísicas, más influyentes para el modelo del reloj interno es la ley de Weber. Según esta ley, existe una relación entre la variabilidad de un juicio temporal y la media del juicio, es decir, entre más aumenta el valor temporal que ha de ser cronometrado, más imprecisas son las estimaciones de la duración de este.
En la actualidad, la utilización de estos métodos de análisis y el incremento de la utilización de técnicas de neuroimagen han permitido a varios modelos cognitivos desarrollar y consolidar la idea de la existencia de ese reloj, proporcionándonos una nueva perspectiva de cómo construimos esa representación subjetiva.
A continuación se presentan dos de los modelos psicológicos del reloj interno más importantes: el modelo del oscilador temporal interno (5) y el modelo de cronometraje escalar (6).
Modelo del oscilador temporal interno (Treisman, 1963)
Este modelo surge después de los trabajos pioneros de François (1927) y Hoagland (1933) en los que reportaron el efecto de la modulación de la temperatura en la estimación de los juicios temporales.
Treisman (1963) retoma y desarrolla esta idea en el primer modelo de procesamiento de la información temporal basado en el postulado del reloj interno (Figura 1).

Según este modelo, el reloj está compuesto de un marcapasos que emite una serie de pulsiones o “clics” con una frecuencia regular, un contador (acumulador) y un interruptor.
Según este modelo, cuando un estímulo es percibido por alguna modalidad sensorial, el interruptor que conecta el marcapasos con el contador se cierra, permitiéndole a las pulsiones (clics) ingresar al contador en donde son acumulados, para luego ser enviadas a un almacén de memoria a corto plazo. Una vez que el estímulo desaparece, el interruptor se abre nuevamente, terminando con la conexión entre el marcapasos y el contador. Según Treisman (1963), la duración del tiempo subjetivo depende del número de pulsiones o “clics” contabilizados en el acumulador. El número de pulsos acumulados durante un determinado intervalo temporal constituye la base por la cual discriminamos las duraciones de los eventos. El nivel de activación del marcapasos puede variar, ya que la velocidad con que el marcapasos produce los pulsos es sensible a los estímulos externos, actuando también como modulador de la velocidad del contador.
Para comprobar estas hipótesis, los investigadores utilizaron un método que consistía en presentar a los participantes una sucesión repetitiva de clics o destellos visuales antes o al mismo tiempo que un estímulo temporal (7). Este método, les permitió mostrar que una frecuencia de clics entre 2.5 y 27.5 Hz puede cambiar la percepción del tiempo, de tal manera que la duración de estímulo temporal se juzga como más larga cuando el estímulo temporal es presentado con clics repetitivos. Además, pudieron observar que la magnitud del efecto aumenta con la frecuencia de los clics (8,9).
Este efecto se ha replicado en numerosos estudios usando diferentes tareas temporales, demostrando así la robustez de su hipótesis. (10–15).
La evidencia más contundente sobre la manera en la que funciona el marcapasos proviene de estudios farmacológicos realizados en el modelo animal (16). Estos estudios demostraron que un agonista de la dopamina, el metilfenidato aumenta la velocidad del ritmo del marcapasos, mientras que un antagonista el haroperidol lo enlentece.
Posteriormente, resultados similares fueron replicados en seres humanos (17) y reportados en estudios clínicos de pacientes que sufren de la enfermedad de Parkinson, cuya percepción del tiempo se ha visto alterada, posiblemente debido a una pérdida de las células dopaminérgicas de la sustancia negra (18). Sin embargo, aunque el ritmo del marcapasos produzca clics a una tasa constante, las diferentes esperas (apertura – cierre) en el funcionamiento del interruptor, introduce variabilidad en el número de clics que son contabilizadas en el contador.
Por consiguiente, las teorías del reloj interno se interesaron en los efectos de la manipulación de la atención sobre la percepción del tiempo, a través de situaciones experimentales que modificaban los tiempos de apertura y cierre del interruptor.
El principal paradigma experimental utilizado para el estudio del efecto de la atención en la estimación del tiempo fue el paradigma de la doble tarea. En estas condiciones, el participante debía realizar de manera simultánea dos tareas, una tarea temporal y una tarea no temporal (19,20). Estos estudios reportaron que entre menos atención utilice el participante para estimar la duración del estímulo, menor o más corto será el juicio emitido del tiempo.
En el contexto de este modelo, un reciente estudio demostró que la atención juega un rol en el funcionamiento del interruptor (21); así pues, cuando menos recursos atencionales son atribuidos al procesamiento temporal, más tiempo dura abierto el interruptor, menos pulsaciones son acumuladas y, por ende, más corto es el cálculo. En otras palabras, una mayor cantidad de atención sería relevante para la estimación correcta del tiempo.
No obstante, y a pesar de que diferentes estudios muestran cómo las diversas variaciones del tiempo subjetivo pueden explicarse por el funcionamiento de un reloj interno, y más precisamente por el funcionamiento de un marcapasos-interruptor, es en el modelo de cronometraje escalar (6) en donde otros componentes, aparte del reloj interno, son propuestos y desarrollados para explicar los procesamientos temporales más complejos.
El modelo de cronometraje escalar (Gibbon, Chuck y Meck, 1984)
Actualmente, éste es uno de los modelos más influyentes para predecir los comportamientos temporales en los humanos (22–24). Según este modelo, la percepción del tiempo es el resultado de un sistema de procesamiento de la información conformado por cinco módulos que están divididos en tres niveles diferentes (Figura. 2).
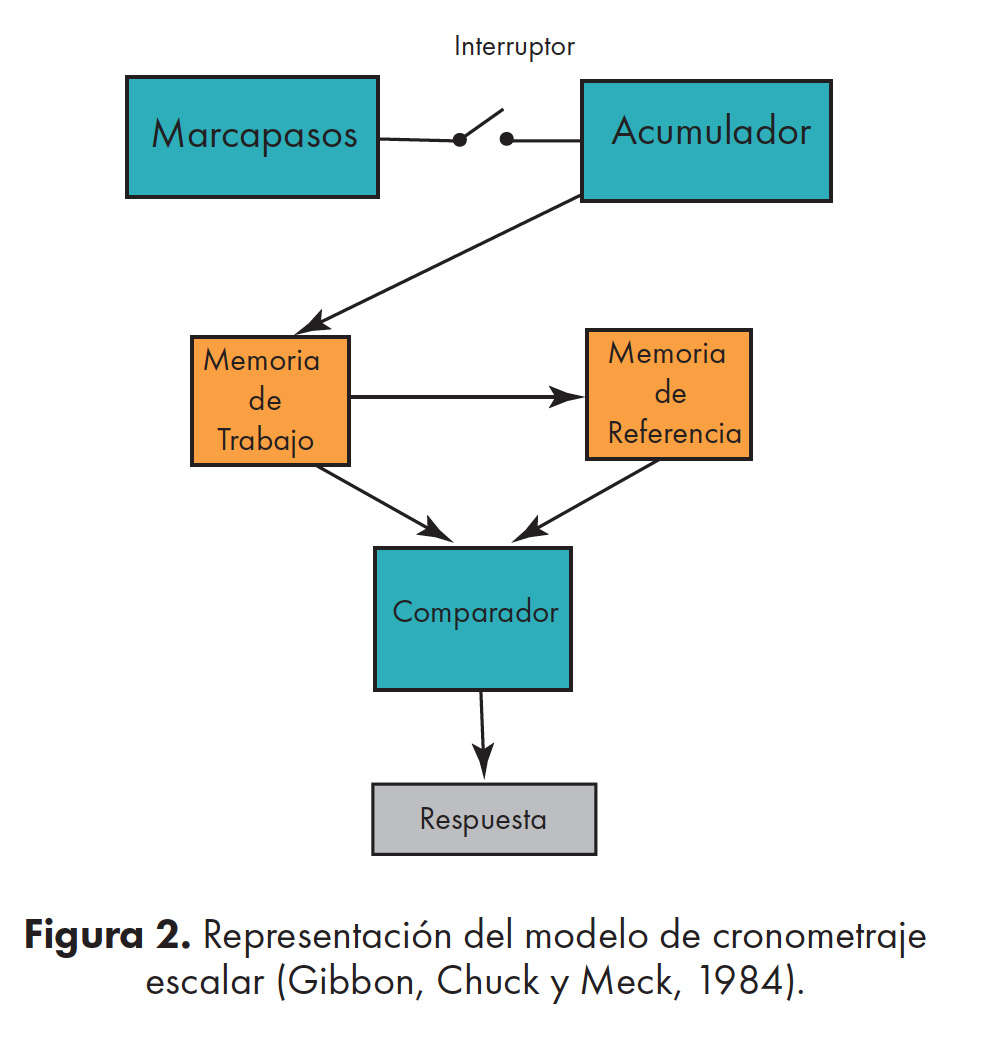
1) Cronometraje (marcapasos y acumulador) 2) almacenamiento (Memoria de trabajo y memoria de referencia) 3) Toma de decisiones.
En un primer nivel, el proceso de cronometraje inicia con el marcapasos o base del tiempo. Este mecanismo interno produce unos pulsos (clics) de manera regular, en un rango que puede oscilar de segundos a minutos. Estos pulsos transitan a través de un interruptor que se cierra hasta llegar al acumulador (anteriormente llamado contador), en donde los pulsos serán registrados y sumados de manera absoluta. Cuando acaba el intervalo de tiempo, los pulsos son transferidos a la memoria de trabajo en el segundo nivel, para ser almacenados y luego comparados con las representaciones temporales anteriormente almacenadas en la memoria de referencia.
La memoria de trabajo guarda la información relativa al ensayo en curso y es únicamente utilizada cuando existe un periodo de tiempo entre el estímulo y la respuesta. Sin embargo, la memoria de referencia almacena de manera permanente la información relacionada con los ensayos anteriores.
En el tercer nivel, correspondiente a la toma de decisión, se establece una comparación del valor del ensayo actual, almacenado en la memoria de trabajo, y el valor almacenado previamente en la memoria de referencia. El organismo emitirá una respuesta, basado en la comparación de esas dos duraciones.
No obstante, la pregunta sobre el sustrato neurobiológico de los mecanismos necesarios a la percepción del tiempo fue inicialmente abordada desde el modelo farmacológico en el animal (6,16) y la patología (18,25), proponiendo los ganglios de la base como área acumuladora de los pulsos: el Núcleo caudado / Putámen y la sustancia negra.
Sin embargo, a partir de los años 90, gracias al auge de las técnicas de neuroimagen, se incrementó rápidamente el conocimiento de la implicación de algunas estructuras corticales y subcorticales en la estimación, percepción y reproducción del tiempo.
En una revisión realizada por Macar y colaboradores (26) en la que incluyó datos obtenidos en trabajos de imagen por resonancia magnética funcional (IRMf), tomografía por emisión de positrones (TEP) y Electroencefalografía (EEG), los autores extendieron el sustrato neurobiológico del reloj interno a una red de estructuras que también están implicadas en el proceso de la atención. Las principales áreas cerebrales relacionadas fueron: la corteza prefrontal dorsolateral (27–31), los ganglios de la base (28–33), el área motora suplementaria (27,28,32,34,35), el cerebelo (28,30,32,33), la corteza del cíngulo anterior (29,31,34,36), y el lóbulo parietal inferior derecho (28,29,37).
Otra revisión realizada por Lewis y Miall (38) concluyó que existe una disociación de estructuras según la escala de duración a evaluar. Así pues, la estimación de duraciones inferiores al segundo implica las siguientes regiones: el área motora suplementaria, la corteza sensoriomotora izquierda, los ganglios de la base, el tálamo, el cerebelo derecho, el giro temporal superior y la corteza premotora lateral. Sin embargo, las estructuras implicadas en la estimación de una duración superior al segundo serían: el cerebelo izquierdo, la corteza prefrontal, los lóbulos parietales bilaterales y la corteza prefrontal dorsolateral.
A pesar de que estos estudios presentan un conjunto de resultados que fomenta el debate sobre los sustratos neurobiológicos de la percepción del tiempo subjetivo, desafortunadamente no proporcionan una medida cuantitativa de la probabilidad de activación de una estructura implicada. Este vacío lo llenaría el trabajo reciente de reciente de Wiener y colaboradores (39), quienes realizaron un metaanálisis utilizando el algoritmo ALE (probabilidad de estimación de activación). Este procedimiento consiste en hacer una modelización de las coordenadas estereotáxicas como una distribución de Gauss y probar la posibilidad de activación de todos los vóxeles, lo que permite entonces un análisis cuantitativo de la duración de activación de estas estructuras.
En este metaanálisis, los estudios fueron separados según dos dimensiones: la duración del estímulo (periodos inferiores y superiores a un segundo) y la naturaleza de la respuesta (motriz o perceptual).
El metaanálisis proporcionó tres resultados principales: en primer lugar, las áreas motoras y la circunvolución frontal inferior derecha aparecieron como las únicas estructuras significativamente activas a través de todas las condiciones de procesamiento temporal. En segundo lugar, los datos demostraron que no hay redes distintas pero que se solapan parcialmente de acuerdo con la duración de tiempo a estimar, menor o mayor que el segundo. Las tareas en las que la duración de tiempo a estimar es inferior al segundo, implican a una red subcortical que involucra los ganglios basales y el cerebelo, mientras que las tareas con duraciones más largas que el segundo activarán estructuras corticales como la corteza prefrontal y el área motora suplementaria. En tercer lugar, la naturaleza motriz o perceptiva de la tarea implica también el rol de diferentes regiones cerebrales. El putamen, está más activo cuando las tareas son de tipo motor y el caudado, cuando son de naturaleza perceptiva.
En otra revisión, Allman y colaboradores (40) reportaron un resumen de los recientes trabajos en neuroimagen que estudian la percepción del tiempo en el modelo animal y en seres humanos. Los resultados también corroboran la implicación de un circuito fronto-estriatal, que sería la base del funcionamiento del reloj interno. En este circuito se destaca el rol de la corteza frontal dorsolateral derecha (41–43) y el del área motora suplementaria y pre-suplementaria, como contador o acumulador del tiempo (44,45).
Estudios que utilizan técnicas muy diferentes proponen que el AMS está relacionada con la función de acumulador del tiempo (34,45–47).
Evidencia de estudios electrofisiológicos.
La variación contingente negativa (en Ingles, Contingent Negative Variation, CNV) es uno de los componentes fisiológicos más asociados con los procesos de estimación temporal. Este es uno de los primeros potenciales de acción evocado, reportado en Electroencefalografía (EEG), cuya naturaleza es principalmente frontal, ha sido interpretado como un indicador de la expectativa de un sujeto ante la aparición de un estímulo precedido por una señal de alerta (48). Algunos estudios reportan una relación entre la latencia de la CNV previa a la respuesta motora y la longitud del intervalo estimado en una tarea de producción de tiempo motor (49), mientras que otros estudios encontraron relación entre la latencia y amplitud de la CNV y la estimación del tiempo utilizando tareas de estimación temporal (46,50–53).
Estudios de electroencefalografía que utilizan métodos de análisis de señal más sofisticados y precisos como el de Laplacian, han reportado también una relación entre la amplitud de la actividad EEG y la duración producida por los sujetos sobre el AMS (46,51).
En resumen, la totalidad de esos estudios sugieren que el AMS podría jugar un rol de acumulador de pulsaciones. Sin embargo, otros estudios sugieren que la variación de la CNV podría, igualmente, estar relacionada con otro componente del reloj, como la memoria o el de la toma de decisiones (54,55).
Evidencia de estudios en el modelo animal.
Estudios realizados en el modelo animal (56) registraron la actividad neuronal del simio despierto mientras realizaba una tarea de percepción o discriminación del tiempo, demostrando que la actividad neuronal de las regiones AMS y pre-AMS se comporta como acumulador, cuando los periodos de preparación a la acción son fijos, es decir, cuando el simio podía predecir la preparación a la acción.
Por su parte, Akkal y colaboradores (57) reportaron resultados que van en el mismo sentido, cuando registraban la actividad unitaria de las neuronas en el mono mientras realizaban una tarea de tiempos de reacción con un periodo preparatorio fijo o variable. En sus estudios, Akkan y colaboradores notaron que las neuronas de la pre-SMA acumulaban la activación, únicamente en el caso del periodo preparatorio fijo. Los autores concluyeron, que ese patrón fisiológico podía representar el sustrato neuronal del acumulador temporal.
Evidencia de estudios en IRMf
Estudios en IRMf también aportan argumentos del rol del AMS como acumulador (58). El trabajo de Coull y Nobre, está inspirado en la idea de Chawla y colaboradores (59) en el cual se observó que la tasa de activación en las áreas visuales aumenta cuando la atención se fija en estímulos visuales.
Inspirados en esta idea, los autores plantearon la hipótesis según la cual, si la atención se dirige hacia el tratamiento del tiempo, las áreas encargadas de este procesamiento deberían activarse. Para este propósito, utilizaron un protocolo de doble tarea, en el cual, los sujetos debían juzgar simultáneamente, la duración de un estímulo visual y su color, repartiendo la atención según las instrucciones del experimentador.
Los resultados revelaron que no solamente la cantidad de atención determina el rendimiento en el juicio temporal, sino que, además, solamente el AMS presentaba una tasa de activación positivamente correlacionada con la cantidad de atención acordada a la duración. Así pues, los autores concluyeron que el AMS se comporta como un acumulador de tiempo. Esa hipótesis fue probada en posteriores estudios (41,60) en los que se demostró que sólo el AMS se compromete en dos fases: codificación y reproducción, pudiendo entonces concluir el rol de esta área en el proceso de acumulación temporal.
Además, el cerebelo lateral izquierdo, podría también participar en ese circuito que integra el AMS. El AMS tiene conexiones robustas con los ganglios basales (61) por lo tanto, podría ser un vínculo entre dos circuitos parcialmente distintos. Esta hipótesis es, por otra parte, totalmente coherente con los resultados de un metaanálisis reciente que tiende a mostrar una disociación funcional entre el pre-AMS y el AMS en el procesamiento de tiempo.
La primera estaría más bien involucrada en un contexto sensorial, con estímulos no secuenciales y en duraciones más largas; mientras que el AMS jugaría un rol más bien cuando el contexto es sensorio motor (62).
Procesamiento temporal y déficit de atención e hiperactividad (TDAH).
Además de la tríada clínica que caracteriza a las personas diagnosticadas con TDAH (inatención, impulsividad e hiperactividad), también se han reportado dificultades en la organización de la conducta (63) Este problema de organización a menudo ha sido considerado como el resultado de una incapacidad para anticipar y predecir eventos, dos habilidades que requieren una percepción exacta de las duraciones temporales.
Se cree que el comportamiento impulsivo en niños TDAH podría ser el resultado de una tendencia a responder y/o actuar demasiado pronto, es decir, con una pobre regulación del tiempo, lo cual podría explicar por qué los pacientes diagnosticados con TDAH tienen dificultad para esperar su turno y dar respuestas oportunas.
Diferencias significativas en el procesamiento temporal de personas diagnosticadas con TDAH han sido principalmente reportadas en niños (64,65) en tareas de percepción y reproducción del tiempo. En las tareas de percepción, los niños con TDAH estiman el tiempo como más largo que sus pares no diagnosticados, aunque en las tareas de reproducción, reproducen el tiempo como más corto.
Por otro lado, según nuestro conocimiento, solo algunos estudios reportan diferencias en adultos TDAH en tareas que estudian del tiempo: en reproducción del tiempo (66) y de bisección temporal.
En un reciente estudio, Suárez y colaboradores (2013) compararon el desempeño de jóvenes adultos diagnosticados con TDAH y de controles sanos, en una tarea de bisección temporal realizada en la modalidad auditiva y visual. Los análisis psicofísicos realizados, utilizando el Difference Limen (DL) y el punto de igualdad subjetivo (PSE), revelaron que en ambas modalidades los sujetos diagnosticados con TDAH aumentaron la variabilidad de los juicios temporales y sobreestimaron las duraciones, en comparación con los controles. Teniendo en cuenta estos resultados, y en el contexto del modelo del reloj interno, los autores sugirieron que estos pacientes pueden tener dificultades para mantener un nivel de atención sostenida o alerta constante, posiblemente debido a una variabilidad en la apertura del interruptor. Además, la tendencia de estos pacientes a sobreestimar el tiempo podría explicarse como dificultades en la capacidad de almacenamiento de pulsaciones en la memoria.
Esta idea de un disfuncionamiento en la variabilidad del interruptor, y por ende en la acumulación de pulsaciones, es coherente con previas hipótesis de modelos neuropsicológicos de la aversión a la espera, reportado anteriormente en el TDAH (70).
Ante la espera de un evento, el paciente con diagnóstico de TDAH, parece tener dificultades para esperar y, en este caso, trataría de reducir el tiempo de espera maximizando su atención en diversas informaciones presentes en el medio ambiente, lo que aceleraría el “paso del tiempo”.
Según este modelo, los niños con TDAH terminarían la tarea más rápido para evitar esperar. La aversión al retraso que, según el modelo propuesto por Sonuga-Barke y colaboradores, caracteriza el TDAH, podría deberse a un disfuncionamiento del reloj interno, que, en este caso, presentaría disfunciones en cuanto a la apertura del interruptor y, por ende, un número desproporcionado de clics acumulados en el contador.
Adicionalmente, estudios de neuroimagen, reportan que pacientes diagnosticados con TDAH presentan diferencias en la activación de regiones cerebrales respecto a sujetos controles sanos, sobre todo en una red estriato-fronto-cerebelosa (71–73), red que también está implicada en el procesamiento del tiempo.
Las fortalezas de los modelos psicológicos del tiempo, junto con el enfoque psicofísico y la comprensión de los mecanismos neurobiológicos, proporcionan en la actualidad un terreno fértil para la investigación del reloj interno. Adicionalmente, este modelo proporciona un contexto prometedor que quizás sea la clave elusiva a la comprensión de los síntomas reportados en personas diagnosticadas con déficit de atención e hiperactividad.
- François M. Contribution à l’étude du sens du temps : la température interne comme facteur de variation de l’appréciation subjective des durées. L’Année psychologique. 27.a ed. 1927;186-204.
- Hoagland H. The Physiological Control of Judgments of Duration: Evidence for a Chemical Clock. The Journal of General Psychology. 1933; 267-87.
- Wearden JH, Penton-Voak IS. Feeling the heat: body temperature and the rate of subjective time, revisited. Q J Exp Psychol B. mayo de 1995;48(2):129-41.
- Block R. Cognitive models of psychological time. 2010.a ed. New York: Psychology Press; 1990.
- Treisman M. Temporal discrimination and the indifference interval. Implications for a model of the «internal clock». Psychol Monogr. 1963;77(13):1-31.
- Gibbon J, Church RM, Meck WH. Scalar timing in memory. Ann N Y Acad Sci. 1984;423:52-77.
- Treisman M, Faulkner A, Naish PL, Brogan D. The internal clock: evidence for a temporal oscillator underlying time perception with some estimates of its characteristic frequency. Perception. 1990;19(6):705-43.
- Treisman M, Brogan D. Time perception and the internal clock: Effects of visual flicker on the temporal oscillator. European Journal of Cognitive Psychology. 4.a ed. 1994;41-70.
- Treisman M, Cook N, Naish PL, MacCrone JK. The internal clock: electroencephalographic evidence for oscillatory processes underlying time perception. Q J Exp Psychol A. mayo de 1994;47(2):241-89.
- Droit-Volet S. Time dilation in children and adults: The idea of a slower internal clock in young children tested with different click frequencies. Behav Processes. mayo de 2017;138:152-9.
- Herbst SK, Javadi AH, van der Meer E, Busch NA. How long depends on how fast–perceived flicker dilates subjective duration. PloS One. 2013;8(10):e76074.
- Jones LA, Allely CS, Wearden JH. Click trains and the rate of information processing: does «speeding up» subjective time make other psychological processes run faster? Q J Exp Psychol 2006. febrero de 2011;64(2):363-80.
- Jones LA, Ogden RS. Vibrotactile timing: Are vibrotactile judgements of duration affected by repetitive stimulation? Q J Exp Psychol 2006. 2016;69(1):75-88.
- Kanai R, Paffen CLE, Hogendoorn H, Verstraten FAJ. Time dilation in dynamic visual display. J Vis. 15 de diciembre de 2006;6(12):1421-30.
- Penton-Voak IS, Edwards H, Percival A, Wearden JH. Speeding up an internal clock in humans? Effects of click trains on subjective duration. J Exp Psychol Anim Behav Process. julio de 1996;22(3):307-20.
- Meck W. Neuropharmacology of timing and time perception. Brain Res Cogn Brain Res. enero de 1998;6(3):233.
- Rammsayer T. Are There Dissociable Roles of the Mesostriatal and Mesolimbocortical Dopamine Systems on Temporal Information Processing in Humans? Neuropsychobiology. 35.a ed. 1997;36-45.
- Artieda J, Pastor MA, Lacruz F, Obeso JA. Temporal discrimination is abnormal in Parkinson’s disease. Brain J Neurol. febrero de 1992;115 Pt 1:199-210.
- Casini L, Macar F. Effects of attention manipulation on judgments of duration and of intensity in the visual modality. Mem Cognit. noviembre de 1997;25(6):812-8.
- Macar F, Grondin S, Casini L. Controlled attention sharing influences time estimation. Mem Cognit. noviembre de 1994;22(6):673-86.
- Burle B, Casini L. Dissociation between activation and attention effects in time estimation: implications for internal clock models. J Exp Psychol Hum Percept Perform. febrero de 2001;27(1):195-205.
- Meck W. Neuropharmacology of timing and time perception. Brain Res Cogn Brain Res. junio de 1996;3(3-4):227-42.
- Meck WH, Penney TB, Pouthas V. Cortico-striatal representation of time in animals and humans. Curr Opin Neurobiol. abril de 2008;18(2):145-52.
- Wearden JH, Philpott K, Win T. Speeding up and (…relatively…) slowing down an internal clock in humans. Behav Processes. 3 de mayo de 1999;46(1):63-73.
- Malapani C, Rakitin B, Levy R, Meck WH, Deweer B, Dubois B, et al. Coupled temporal memories in Parkinson’s disease: a dopamine-related dysfunction. J Cogn Neurosci. mayo de 1998;10(3):316-31.
- Macar F, Lejeune H, Bonnet M, Ferrara A, Pouthas V, Vidal F, et al. Activation of the supplementary motor area and of attentional networks during temporal processing. Exp Brain Res. febrero de 2002;142(4):475-85.
- Brunia CH, de Jong BM, van den Berg-Lenssen MM, Paans AM. Visual feedback about time estimation is related to a right hemisphere activation measured by PET. Exp Brain Res. febrero de 2000;130(3):328-37.
- Jäncke L, Loose R, Lutz K, Specht K, Shah NJ. Cortical activations during paced finger-tapping applying visual and auditory pacing stimuli. Brain Res Cogn Brain Res. septiembre de 2000;10(1-2):51-66.
- Lejeune H, Maquet P, Bonnet M, Casini L, Ferrara A, Macar F, et al. The basic pattern of activation in motor and sensory temporal tasks: positron emission tomography data. Neurosci Lett. 10 de octubre de 1997;235(1-2):21-4.
- Rao SM, Mayer AR, Harrington DL. The evolution of brain activation during temporal processing. Nat Neurosci. marzo de 2001;4(3):317-23.
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams S, Simmons A, et al. Prefrontal involvement in «temporal bridging» and timing movement. Neuropsychologia. diciembre de 1998;36(12):1283-93.
- Jueptner M, Rijntjes M, Weiller C, Faiss JH, Timmann D, Mueller SP, et al. Localization of a cerebellar timing process using PET. Neurology. agosto de 1995;45(8):1540-5.
- Schubotz RI, Friederici AD, von Cramon DY. Time perception and motor timing: a common cortical and subcortical basis revealed by fMRI. NeuroImage. enero de 2000;11(1):1-12.
- Casini L, Vidal F. The SMAs: Neural Substrate of the Temporal Accumulator? Front Integr Neurosci. 2011;5:35.
- Vidal F, Bonnet M, Macar F. Programming the duration of a motor sequence: role of the primary and supplementary motor areas in man. Exp Brain Res. 1995;106(2):339-50.
- Kawashima R, Inoue K, Sugiura M, Okada K, Ogawa A, Fukuda H. A positron emission tomography study of self-paced finger movements at different frequencies. Neuroscience. 1999;92(1):107-12.
- Maquet P, Lejeune H, Pouthas V, Bonnet M, Casini L, Macar F, et al. Brain activation induced by estimation of duration: a PET study. NeuroImage. abril de 1996;3(2):119-26.
- Lewis PA, Miall RC. A right hemispheric prefrontal system for cognitive time measurement. Behav Processes. 28 de febrero de 2006;71(2-3):226-34.
- Wiener M, Hamilton R, Turkeltaub P, Matell MS, Coslett HB. Fast forward: supramarginal gyrus stimulation alters time measurement. J Cogn Neurosci. enero de 2010;22(1):23-31.
- Allman MJ, Teki S, Griffiths TD, Meck WH. Properties of the internal clock: first- and second-order principles of subjective time. Annu Rev Psychol. 2014;65:743-71.
- Coull JT, Nazarian B, Vidal F. Timing, storage, and comparison of stimulus duration engage discrete anatomical components of a perceptual timing network. J Cogn Neurosci. diciembre de 2008;20(12):2185-97.
- Harrington DL, Haaland KY, Knight RT. Cortical networks underlying mechanisms of time perception. J Neurosci Off J Soc Neurosci. 1 de febrero de 1998;18(3):1085-95.
- Koch G, Oliveri M, Caltagirone C. Neural networks engaged in milliseconds and seconds time processing: evidence from transcranial magnetic stimulation and patients with cortical or subcortical dysfunction. Philos Trans R Soc Lond B Biol Sci. 12 de julio de 2009;364(1525):1907-18.
- Grahn JA, McAuley JD. Neural bases of individual differences in beat perception. NeuroImage. 1 de octubre de 2009;47(4):1894-903.
- Wencil EB, Coslett HB, Aguirre GK, Chatterjee A. Carving the clock at its component joints: neural bases for interval timing. J Neurophysiol. julio de 2010;104(1):160-8.
- Macar F, Vidal F, Casini L. The supplementary motor area in motor and sensory timing: evidence from slow brain potential changes. Exp Brain Res. abril de 1999;125(3):271-80.
- Macar F. Expectancy, controlled attention and automatic attention in prospective temporal judgments. Acta Psychol (Amst). septiembre de 2002;111(2):243-62.
- Walter WG, Cooper R, Aldridge VJ, Mccallum WC, Winter AL. Contingent negative variation: an electric sign of sensorimotor association and expectancy in the human brain. Nature. 25 de julio de 1964;203:380-4.
- Ruchkin DS, McCalley MG, Glaser EM. Event related potentials and time estimation. Psychophysiology. septiembre de 1977;14(5):451-5.
- Ghadirian AM, Dubrovsky B. A longitudinal CNV study of the evolution and treatment of bipolar illness. Int J Neurosci. octubre de 1993;72(3-4):245-50.
- Macar F, Vidal F. The CNV peak: an index of decision making and temporal memory. Psychophysiology. noviembre de 2003;40(6):950-4.
- Pfeuty M, Ragot R, Pouthas V. Processes involved in tempo perception: a CNV analysis. Psychophysiology. enero de 2003;40(1):69-76.
- Pfeuty M, Ragot R, Pouthas V. Relationship between CNV and timing of an upcoming event. Neurosci Lett. 1 de julio de 2005;382(1-2):106-11.
- Pouthas V, George N, Poline J-B, Pfeuty M, Vandemoorteele P-F, Hugueville L, et al. Neural network involved in time perception: an fMRI study comparing long and short interval estimation. Hum Brain Mapp. agosto de 2005;25(4):433-41.
- van Rijn H, Kononowicz TW, Meck WH, Ng KK, Penney TB. Contingent negative variation and its relation to time estimation: a theoretical evaluation. Front Integr Neurosci. 2011;5:91.
- Mita A, Mushiake H, Shima K, Matsuzaka Y, Tanji J. Interval time coding by neurons in the presupplementary and supplementary motor areas. Nat Neurosci. abril de 2009;12(4):502-7.
- Akkal D, Escola L, Bioulac B, Burbaud P. Time predictability modulates pre-supplementary motor area neuronal activity. Neuroreport. 7 de junio de 2004;15(8):1283-6.
- Coull J, Nobre A. Dissociating explicit timing from temporal expectation with fMRI. Curr Opin Neurobiol. abril de 2008;18(2):137-44.
- Chawla D, Rees G, Friston KJ. The physiological basis of attentional modulation in extrastriate visual areas. Nat Neurosci. julio de 1999;2(7):671-6.
- Coull JT, Cheng R-K, Meck WH. Neuroanatomical and neurochemical substrates of timing. Neuropsychopharmacol Off Publ Am Coll Neuropsychopharmacol. enero de 2011;36(1):3-25.
- Akkal D, Dum RP, Strick PL. Supplementary motor area and presupplementary motor area: targets of basal ganglia and cerebellar output. J Neurosci Off J Soc Neurosci. 3 de octubre de 2007;27(40):10659-73.
- Schwartze M, Rothermich K, Kotz SA. Functional dissociation of pre-SMA and SMA-proper in temporal processing. NeuroImage. marzo de 2012;60(1):290-8.
- Barkley RA. Behavioral inhibition, sustained attention, and executive functions: Constructing a unifying theory of ADHD. Psychol Bull. enero de 1997;121(1):65-94.
- Toplak ME, Dockstader C, Tannock R. Temporal information processing in ADHD: findings to date and new methods. J Neurosci Methods. 15 de febrero de 2006;151(1):15-29.
- Toplak ME, Tannock R. Time Perception: Modality and Duration Effects in Attention-Deficit/Hyperactivity Disorder (ADHD). J Abnorm Child Psychol. octubre de 2005;33(5):639-54.
- Barkley RA, Murphy KR, Bush T. Time perception and reproduction in young adults with attention deficit hyperactivity disorder. Neuropsychology. julio de 2001;15(3):351-60.
- Pollak Y, Kroyzer N, Yakir A, Friedler M. Testing possible mechanisms of deficient supra-second time estimation in adults with attention-deficit/hyperactivity disorder. Neuropsychology. septiembre de 2009;23(5):679-86.
- Valera EM, Spencer RMC, Zeffiro TA, Makris N, Spencer TJ, Faraone SV, et al. Neural substrates of impaired sensorimotor timing in adult attention-deficit/hyperactivity disorder. Biol Psychiatry. 15 de agosto de 2010;68(4):359-67.
- Suarez I, Lopera F, Pineda D, Casini L. The cognitive structure of time estimation impairments in adults with attention deficit hyperactivity disorder. Cogn Neuropsychol. 2013;30(4):195-207.
- Sonuga-Barke EJ, Taylor E, Sembi S, Smith J. Hyperactivity and delay aversion–I. The effect of delay on choice. J Child Psychol Psychiatry. febrero de 1992;33(2):387-98.
- Cubillo A, Halari R, Ecker C, Giampietro V, Taylor E, Rubia K. Reduced Activation and Inter-Regional Functional Connectivity of Fronto-Striatal Networks in Adults with Childhood Attention-Deficit Hyperactivity Disorder (adhd) and Persisting Symptoms During Tasks of Motor Inhibition and Cognitive Switching. J Psychiatr Res. juillet de 2010;44(10):629-39.
- Noreika V, Falter CM, Rubia K. Timing deficits in attention-deficit/hyperactivity disorder (ADHD): evidence from neurocognitive and neuroimaging studies. Neuropsychologia. enero de 2013;51(2):235-66.
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. Hypofrontality in attention deficit hyperactivity disorder during higher-order motor control: a study with functional MRI. Am J Psychiatry. junio de 1999;156(6):891-6.